В 1863 году И.М. Сеченов открыл процесс торможения в ЦНС.
Торможение существует наряду с возбуждением и представляет собой одну из форм деятельности нейрона. Торможением называют особый нервный процесс, выражающийся в уменьшении или полном отсутствии ответной реакции на раздражение.
Начало изучения торможения в центральной нервной системе связывают с выходом в свет работы И.М.Сеченого «Рефлексы головного мозга» (1863), в которой он показал возможность торможения двигательных рефлексов лягушки при химическом раздражении зрительных бугров головного мозга.
Классический опыт Сеченова заключается в следующем: у лягушки с перерезанным головным мозгом на уровне зрительных бугров определяла время сгибательного рефлекса при раздражении лапки серной кислотой. После этого на зрительные бугры накладывали кристаллик поваренной соли и снова определяли время рефлекса. Оно постепенно увеличивалось, вплоть до полного исчезновения реакции. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса постепенно восстанавливалось. Это позволило говорить о том, что торможение – активный процесс, возникающий при раздражении определенных отделов центральной нервной системы.
Позже И.М.Сеченовым и его учениками было показано, что торможение в центральной нервной системе может возникнуть при нанесении сильного раздражения на любые афферентные пути.
Виды и механизмы торможения. Благодаря микроэлектродной технике исследования стало возможным изучение процесса торможения на клеточном уровне.
В центральной нервной системе наряду с возбуждающими имеются и тормозящие нейроны. На каждой нервной клетке располагаются возбуждающие и тормозящие синапсы. А поэтому в каждый данный момент на теле нейрона возникает в одних синапсах возбуждение, а в других – торможение; соотношение этих процессов определяет характер ответной реакции.
Различают два вида торможения в зависимости от механизмов его возникновения: деполяризационное гиперполяризационное. Деполяризационное торможение возникает вследствие длительной деполяризации мембраны, а гиперполяризационное – вследствие гиперполяризации мембраны.
Наступлению деполяризационного торможения предшествует состояние возбуждения. Вследствие длительного раздражения это возбуждение переходит в торможение. В основе возникновения деполяризационного торможения лежит инактивация мембраны по натрию, вследствие сего уменьшается потенциал действия и его раздражающее влияние на соседние участки, в итоге прекращается проведение возбуждения.
Гиперполяризационное торможение осуществляется с участием особых тормозных структур и связано с изменением проницаемости мембраны по отношению к калию и хлору, что вызывает увеличение мембранного и порогового потенциалов, в результате чего становится невозможной ответная реакция.
По характеру возникновения различают первичное и вторичное торможение. Первичное торможение возникает под влиянием раздражения сразу без предварительного возбуждения и осуществляется с участием тормозных синапсов. Вторичное торможение осуществляется без участия тормозных структур и возникает вследствие перехода возбуждения в торможение.
Первичное торможение по механизму возникновения может быть гиперполяризационным и деполяризационным, а по месту возникновения – постсинаптическим и пресинаптическим.
Первичное гиперполяризационное постсинаптическое торможение характерно для мотонейронов и осуществляется через вставочный тормозной нейрон. Импульс, пришедший к тормозному синапсу, вызывает гиперполяризацию постсинаптической мембраны мотонейрона. При этом возрастает величина МП на 5-8 мВ. Это увеличение МП называют тормозным постсинаптическим потенциалом (ТПСП). Величина и длительность тормозного постсинаптического потенциала зависят от силы раздражения и его взаимодействия с возбуждающим постсинаптическим потенциалом (ВПСП).
Постсинаптическое торможение связано с выделением в синапсах медиатора, который изменяет ионную проницаемость постсинаптической мембраны. Хорошо изучено открытое Экклосом и сотрудниками (1954) постсинаптическое торможение мотонейрона, возникающее под влиянием клеток Реншоу. Клетки Реншоу располагаются в передних рогах спинного мозга и обладают высокой электрической активностью. Они могут даже в ответ на одиночный пресинаптический импульс генерировать потенциалы очень высокой частоты – до 1400 импульсов в секунду. Возбуждение к клеткам Реншоу приходит антидромно (в обратном направлении) по разветвлениям аксона мотонейрона, отходящим от него при выходе из спинного мозга. В свою очередь аксон клетки Реншоу контактирует с сомой этого же мотонейрона. Возбуждение, пришедшее антидромно к клетке Реншоу, вызывает в ней высокочастотный разряд, под влиянием которого в мотонейроне создается ТПСП, длящийся до 100 мс. Этот вид постсинаптического торможения называют возвратным или антидромным торможением. Медиатор клетки Реншоу является ацетилхолин.
Первичное деполяризационное пресинаптическое торможение
Развивается в пресинаптических разветвлениях аксонов афферентных нейронов, к которым подходят окончания промежуточных нейронов, образующие на них аксональные синапсы. Эти нейроны обладают высокой электрической активностью. Посылая высокочастотные разряды, они создают на пресинаптических разветвлениях афферентных аксонов длительную деполяризацию (до нескольких сотен миллисекунд). В связи с этим здесь блокируется проведение импульсов, идущих к синапсам мотонейронов, вследствие чего уменьшается или полностью прекращается их активность.
Пресинаптическое торможение является широко распространенным механизмом в ЦНС. Установлено, что оно может быть вызвано не только импульсами с афферентным волокном, но и при раздражении различных структур головного мозга.
Вторичное торможение осуществляется без участия специальных тормозных структур и развивается в возбуждающих синапсах. Такого типа торможение было изучено Н.Е.Введенским (1886) и названо пессимальным торможением в любом участке, обладающем низкой лабильностью (например, в нервно-мышечном синапсе или в синапсах ЦНС). По механизму возникновения вторичное торможение может быть деполяризационным и гиперполяризационным. Вторичным деполяризационным торможением являются рефрактерность и пессимальное торможение.
Механизм возникновения пессимального торможения детально изучен на нервно-мышечных синапсах. Установлено, что в основе его развития лежит стойкая деполяризация, которая может возникнуть как в постсинаптической, так и в пресинаптической мембране синапса под влиянием частой стимуляции.
Вторичное гиперполяризационное торможение возникает после возбуждения в тех же самых нейронах. При сильном возбуждении нейронов их ПД сопровождается последующей длительной гиперполяризацией, наступающей вследствие повышения проницаемости мембраны по калию. Поэтому возникающий при данной силе раздражения ВПСП становится недостаточным для того, что бы деполяризовать мембрану до критического уровня. В результате наблюдается уменьшение или отсутствие реакции.
Роль торможения.
a. Охранительная роль – для предотвращения истощения медиаторов и прекращения деятельности ЦНС.
b. Участвует в обработке поступающей в ЦНС информации.
c. Торможение важный фактор обеспечения координационной деятельности ЦНС.
15. Координационная деятельность ЦНС. Механизмы координации. Факторы, обеспечивающие возможность координации.
Понятие о координации. Приспособление организма к различным изменениям внешней среды возможно благодаря наличию в ЦНС координации функций. Под координацией понимают взаимодействие нейронов, а, следовательно, и нервных процессов, в ЦНС, которое обеспечивает ее согласованную деятельность, направленную на интеграцию (объединение) функций различных органов и систем организма.
Известен ряд механизмов, лежащих в основе координирующей деятельности нервной системы. Одни из них связаны с морфологическими особенностями ее строения (принцип общего конечного пути, принцип обратных связей), другие – с функциональными свойствами (иррадиация, индукция и др.)
Иррадиация возбуждения в центральной нервной системе. В 1908 г. А. А. Ухтомский и Н. Е. Введенский в совместной работе установили, что любое возбуждение, возникающее при раздражении того или иного рецептора, придя в центральную нервную систему, широко по ней распространяется- иррадиирует . Оно захватывает не только центры данного рефлекса, но и другие участки центральной нервной системы. Иррадиация тем шире, чем сильнее и длительное афферентное раздражение.
В основе иррадиации лежат многочисленные связи аксонов афферентных нейронов с дендритами и телами нейронов ЦНС, имеющих большое число контактов с различными нервными центрами и друг с другом. Возбуждение может распространяться на большие расстояния: от нейронов спинного мозга к различным отделам головного мозга вплоть до коры больших полушарий.
Получены экспериментальные данные, позволяющие говорить о закономерностях иррадиации. Оказалось, что в реакцию вовлекаются прежде всего, нейроны, имеющие самый маленький пороговый потенциал, т.е. обладающие наиболее высокой возбудимостью. В них, прежде всего деполяризация достигает критического уровня и возникает волна возбуждения. При увеличении интенсивности раздражения в реакцию вовлекаются менее возбудимые нейроны, при этом процесс возбуждения захватывает все большее количество клеток ЦНС.
Но, несмотря на широкую связь нервных центров, иррадиация возбуждения в ЦНС имеет свои пределы, вследствие чего в деятельное состояние приходят лишь определенные ее отделы.
Процессы индукции в ЦНС. Индукция – один из важнейших принципов координации, который состоит в том, что при возникновении возбуждения в одном из участков ЦНС в сопряженных центрах возникает противоположный процесс – торможение. И, наоборот, при возникновении торможения в одних центрах в сопряженных возникает возбуждение. Индукция ограничивает процесс иррадиации.
Различают одновременную (или пространственную) и последовательную индукцию. При одновременной индукции в одно и то же время в одном центре возникает процесс возбуждения, а в сопряженном центре – торможение (или наоборот). Примером одновременной индукции может быть рассмотренная выше реципрокная иннервация мышц-антагонистов.
Процессам, происходящим в ЦНС, свойственна большая подвижность, без которой невозможно осуществление сложных и быстрых двигательных актов и других ответных реакций. В одном и том же центре осуществляется смена происходящих в нем процессов на противоположные. Смену возбуждения называют отрицательной последовательной индукцией, а торможение на возбуждение – положительной последовательной индукцией . Благодаря такой последовательной смене процессов в нервных центрах возможно чередование сгибательных и разгибательных реакций конечностей, что необходимо для осуществления двигательного акта.
Конвергенция. Импульсы приходящие в ЦНС по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же промежуточным и эфекторным нейронам. Этот факт лег в основу принципа конвергенции, установленного Ч.Шеррингтоном. Конвергенция нервных импульсов объясняется тем, что на теле и дендритах каждого нейрона в ЦНС оканчиваются аксоны множества других нервных клеток. В спинном и продолговатом мозгу конвергенция имеет сравнительно ограниченный характер: на вставочных и моторных нейронах конвергируют афферентные импульсы, возникающие в различных участках рецептивного поля только одного и того же рефлекса. В отличие от этого в высших отделах ЦНС – в подкорковых ядрах и в коре больших полушарий – наблюдается конвергенция импульсов, исходящих из разных рецепторных зон. Поэтому один и тот же нейрон может возбуждаться импульсами, возникающими при раздражении и слуховых, и зрительных, и кожных рецепторов.
Принцип общего конечного пути. Этот принцип исходит из анатомического соотношения между афферентными и эфферентными нейронами. Количество чувствующих нейронов, приносящих возбуждение ЦНС, в 5 раз больше, чем двигательных. Соотношение между ними будет еще больше, если учесть, что вставочные нейроны являются воспринимающими нейронами в ЦНС. В связи с этим к одному мотонейрону приходит множество импульсов от различных рецепторов, но только некоторые из них приобретают рабочее значение. Таким образом, самые разнообразные стимулы могут быть причиной одной и той же рефлекторной реакции, т.е. происходит борьба за «общий конечный путь». Позднее было показано, что не количественное соотношение путей, а функциональные особенности нервных центров определяют, какой из множества нервных импульсов, сталкивающихся на пути к мотонейрону, окажется победителем и завладеет общим конечным путем. В ответ на множество различных раздражений всегда возникает биологически более значимая для организма реакция.
Принцип обратной связи. Воздействие работающего органа на состояние своего центра получило название обратной связи . Она обеспечивает длительное поддержание активности нервных центров, движение процессов возбуждения, торможения в ЦНС и зависит от постоянного притока вторичных афферентных импульсов. Импульсы, которые возникают в результате деятельности различных органов и тканей, называют, вторичными афферентными импульсами , а импульсы, идущие от рецепторов и вызывающие первичный рефлекторный акт, - первичными рефлекторными импульсами.
Вторичные афферентные импульсы возникают в мышцах, сухожилиях и суставах при осуществлении их деятельности. Они, постоянно поступая ото всех органов тела в ЦНС, способствуют ощущению положения нашего тела без зрительного контроля, обеспечивают поддержание нужного уровня функционирования нейронов в каждый данный момент.
Вторичная афферентная импульсация вносит постоянные поправки в осуществляющийся рефлекторный акт и обеспечивает наиболее тонкое приспособление организма к внешним воздействиям.
Афферентные импульсы, идущие от рабочих органов, способствуют созданию аутогенного (собственного) торможения . Оно возникает в результате поступления в ЦНС афферентных импульсов от рецепторов – сухожильных рецепторов Гольджи. Эти рецепторы приходят в состояние возбуждения при растяжении или сокращении мышц. Возникший ТПСП уменьшает степень активности данного мотонейрона. Величина этих изменений может быть различной. Аутогенное торможение обеспечивает лучшее приспособление мышцы к осуществлению рефлекторного двигательного акта.
Факторы, обеспечивающие возможность координации:
1) Фактор структурно-функциональной связи – это наличие между отделами ЦНС, между ЦНС и различными органами функциональной связи, обеспечивающей преимущественное распространение возбуждения между ними. Прямая связь – управление другим центром или рабочим органом с помощью посылки к ни эфферентных импульсов, ПР: мозжечек посылает импульсы к ядрам ствола мозга. Обратная связь (обратная афферентация ) – управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них. Реципрокная связь – обеспечивает торможение центра – антагониста при возбуждении центра-агониста (мышцы сгибатели и разгибатели).
2) Фактор субординации – подчинение нижележащих отделов ЦНС вышележащим.
3) Фактор силы. Принцип общего конечного пути – в борьбе за общий конечный путь побеждает более сильное возбуждение (более важная команда в биологическом отношении), ПР: при слабом раздражении – рефлекс почесывания, при сильном – оборонительный рефлекс сгибание конечности, при одновременном раздражении возникает только оборонительный рефлекс).
4) Одностороннее проведение возбуждения в химических синапсах упорядочивает распространение возбуждения.
5) Феномен облегчения участвует при выработке навыков – возбуждение быстрее распространяется по проторенным путям, навыки становятся более координированными, ненужные движения постепенно устраняются.
6) Доминанта играет важную роль в процессах координации. Обеспечивает автоматизированное выполнение двигательных актов в процессе трудовой деятельности (доминанта двигательных центров).
Непрерывная смена процессов возбуждения и торможения в корковых клетках определяет цикличность работы отдельных органов и всего организма в целом. Этим объясняется иногда кажущаяся невероятная работоспособность некоторых выдающихся людей; недаром говорят, что гениальность на 90% заключается в высокой трудоспособности, которая во многом зависит от рациональной системы работы. Такую глубоко продуманную систему, как правило, создавали для себя все выдающиеся люди.
В 1863 году И.М. Сеченов открыл процесс торможения в ЦНС.
Торможение существует наряду с возбуждением и представляет собой одну из форм деятельности нейрона. Торможением называют особый нервный процесс, выражающийся в уменьшении или полном отсутствии ответной реакции на раздражение.
Начало изучения торможения в центральной нервной системе связывают с выходом в свет работы И.М.Сеченого «Рефлексы головного мозга» (1863), в которой он показал возможность торможения двигательных рефлексов лягушки при химическом раздражении зрительных бугров головного мозга.
Классический опыт Сеченова заключается в следующем: у лягушки с перерезанным головным мозгом на уровне зрительных бугров определяла время сгибательного рефлекса при раздражении лапки серной кислотой. После этого на зрительные бугры накладывали кристаллик поваренной соли и снова определяли время рефлекса. Оно постепенно увеличивалось, вплоть до полного исчезновения реакции. После снятия кристаллика соли и промывания мозга физиологическим раствором время рефлекса постепенно восстанавливалось. Это позволило говорить о том, что торможение – активный процесс, возникающий при раздражении определенных отделов центральной нервной системы.
Позже И.М.Сеченовым и его учениками было показано, что торможение в центральной нервной системе может возникнуть при нанесении сильного раздражения на любые афферентные пути.
Виды и механизмы торможения. Благодаря микроэлектродной технике исследования стало возможным изучение процесса торможения на клеточном уровне.
В центральной нервной системе наряду с возбуждающими имеются и тормозящие нейроны. На каждой нервной клетке располагаются возбуждающие и тормозящие синапсы. А поэтому в каждый данный момент на теле нейрона возникает в одних синапсах возбуждение, а в других – торможение; соотношение этих процессов определяет характер ответной реакции.
Различают два вида торможения в зависимости от механизмов его возникновения: деполяризационное гиперполяризационное. Деполяризационное торможение возникает вследствие длительной деполяризации мембраны, а гиперполяризационное – вследствие гиперполяризации мембраны.
Наступлению деполяризационного торможения предшествует состояние возбуждения. Вследствие длительного раздражения это возбуждение переходит в торможение. В основе возникновения деполяризационного торможения лежит инактивация мембраны по натрию, вследствие сего уменьшается потенциал действия и его раздражающее влияние на соседние участки, в итоге прекращается проведение возбуждения.
Гиперполяризационное торможение осуществляется с участием особых тормозных структур и связано с изменением проницаемости мембраны по отношению к калию и хлору, что вызывает увеличение мембранного и порогового потенциалов, в результате чего становится невозможной ответная реакция.
По характеру возникновения различают первичное и вторичное торможение. Первичное торможение возникает под влиянием раздражения сразу без предварительного возбуждения и осуществляется с участием тормозных синапсов. Вторичное торможение осуществляется без участия тормозных структур и возникает вследствие перехода возбуждения в торможение.
Первичное торможение по механизму возникновения может быть гиперполяризационным и деполяризационным, а по месту возникновения – постсинаптическим и пресинаптическим.
Первичное гиперполяризационное постсинаптическое торможение характерно для мотонейронов и осуществляется через вставочный тормозной нейрон. Импульс, пришедший к тормозному синапсу, вызывает гиперполяризацию постсинаптической мембраны мотонейрона. При этом возрастает величина МП на 5-8 мВ. Это увеличение МП называют тормозным постсинаптическим потенциалом (ТПСП). Величина и длительность тормозного постсинаптического потенциала зависят от силы раздражения и его взаимодействия с возбуждающим постсинаптическим потенциалом (ВПСП).
Постсинаптическое торможение связано с выделением в синапсах медиатора, который изменяет ионную проницаемость постсинаптической мембраны. Хорошо изучено открытое Экклосом и сотрудниками (1954) постсинаптическое торможение мотонейрона, возникающее под влиянием клеток Реншоу. Клетки Реншоу располагаются в передних рогах спинного мозга и обладают высокой электрической активностью. Они могут даже в ответ на одиночный пресинаптический импульс генерировать потенциалы очень высокой частоты – до 1400 импульсов в секунду. Возбуждение к клеткам Реншоу приходит антидромно (в обратном направлении) по разветвлениям аксона мотонейрона, отходящим от него при выходе из спинного мозга. В свою очередь аксон клетки Реншоу контактирует с сомой этого же мотонейрона. Возбуждение, пришедшее антидромно к клетке Реншоу, вызывает в ней высокочастотный разряд, под влиянием которого в мотонейроне создается ТПСП, длящийся до 100 мс. Этот вид постсинаптического торможения называют возвратным или антидромным торможением. Медиатор клетки Реншоу является ацетилхолин.
Первичное деполяризационное пресинаптическое торможение
Развивается в пресинаптических разветвлениях аксонов афферентных нейронов, к которым подходят окончания промежуточных нейронов, образующие на них аксональные синапсы. Эти нейроны обладают высокой электрической активностью. Посылая высокочастотные разряды, они создают на пресинаптических разветвлениях афферентных аксонов длительную деполяризацию (до нескольких сотен миллисекунд). В связи с этим здесь блокируется проведение импульсов, идущих к синапсам мотонейронов, вследствие чего уменьшается или полностью прекращается их активность.
Пресинаптическое торможение является широко распространенным механизмом в ЦНС. Установлено, что оно может быть вызвано не только импульсами с афферентным волокном, но и при раздражении различных структур головного мозга.
Вторичное торможение осуществляется без участия специальных тормозных структур и развивается в возбуждающих синапсах. Такого типа торможение было изучено Н.Е.Введенским (1886) и названо пессимальным торможением в любом участке, обладающем низкой лабильностью (например, в нервно-мышечном синапсе или в синапсах ЦНС). По механизму возникновения вторичное торможение может быть деполяризационным и гиперполяризационным. Вторичным деполяризационным торможением являются рефрактерность и пессимальное торможение.
Механизм возникновения пессимального торможения детально изучен на нервно-мышечных синапсах. Установлено, что в основе его развития лежит стойкая деполяризация, которая может возникнуть как в постсинаптической, так и в пресинаптической мембране синапса под влиянием частой стимуляции.
Вторичное гиперполяризационное торможение возникает после возбуждения в тех же самых нейронах. При сильном возбуждении нейронов их ПД сопровождается последующей длительной гиперполяризацией, наступающей вследствие повышения проницаемости мембраны по калию. Поэтому возникающий при данной силе раздражения ВПСП становится недостаточным для того, что бы деполяризовать мембрану до критического уровня. В результате наблюдается уменьшение или отсутствие реакции.
Роль торможения.
a. Охранительная роль – для предотвращения истощения медиаторов и прекращения деятельности ЦНС.
b. Участвует в обработке поступающей в ЦНС информации.
c. Торможение важный фактор обеспечения координационной деятельности ЦНС.
15. Координационная деятельность ЦНС. Механизмы координации. Факторы, обеспечивающие возможность координации.
Понятие о координации. Приспособление организма к различным изменениям внешней среды возможно благодаря наличию в ЦНС координации функций. Под координацией понимают взаимодействие нейронов, а, следовательно, и нервных процессов, в ЦНС, которое обеспечивает ее согласованную деятельность, направленную на интеграцию (объединение) функций различных органов и систем организма.
Известен ряд механизмов, лежащих в основе координирующей деятельности нервной системы. Одни из них связаны с морфологическими особенностями ее строения (принцип общего конечного пути, принцип обратных связей), другие – с функциональными свойствами (иррадиация, индукция и др.)
Иррадиация возбуждения в центральной нервной системе. В 1908 г. А. А. Ухтомский и Н. Е. Введенский в совместной работе установили, что любое возбуждение, возникающее при раздражении того или иного рецептора, придя в центральную нервную систему, широко по ней распространяется- иррадиирует . Оно захватывает не только центры данного рефлекса, но и другие участки центральной нервной системы. Иррадиация тем шире, чем сильнее и длительное афферентное раздражение.
В основе иррадиации лежат многочисленные связи аксонов афферентных нейронов с дендритами и телами нейронов ЦНС, имеющих большое число контактов с различными нервными центрами и друг с другом. Возбуждение может распространяться на большие расстояния: от нейронов спинного мозга к различным отделам головного мозга вплоть до коры больших полушарий.
Получены экспериментальные данные, позволяющие говорить о закономерностях иррадиации. Оказалось, что в реакцию вовлекаются прежде всего, нейроны, имеющие самый маленький пороговый потенциал, т.е. обладающие наиболее высокой возбудимостью. В них, прежде всего деполяризация достигает критического уровня и возникает волна возбуждения. При увеличении интенсивности раздражения в реакцию вовлекаются менее возбудимые нейроны, при этом процесс возбуждения захватывает все большее количество клеток ЦНС.
Но, несмотря на широкую связь нервных центров, иррадиация возбуждения в ЦНС имеет свои пределы, вследствие чего в деятельное состояние приходят лишь определенные ее отделы.
Процессы индукции в ЦНС. Индукция – один из важнейших принципов координации, который состоит в том, что при возникновении возбуждения в одном из участков ЦНС в сопряженных центрах возникает противоположный процесс – торможение. И, наоборот, при возникновении торможения в одних центрах в сопряженных возникает возбуждение. Индукция ограничивает процесс иррадиации.
Различают одновременную (или пространственную) и последовательную индукцию. При одновременной индукции в одно и то же время в одном центре возникает процесс возбуждения, а в сопряженном центре – торможение (или наоборот). Примером одновременной индукции может быть рассмотренная выше реципрокная иннервация мышц-антагонистов.
Процессам, происходящим в ЦНС, свойственна большая подвижность, без которой невозможно осуществление сложных и быстрых двигательных актов и других ответных реакций. В одном и том же центре осуществляется смена происходящих в нем процессов на противоположные. Смену возбуждения называют отрицательной последовательной индукцией, а торможение на возбуждение – положительной последовательной индукцией . Благодаря такой последовательной смене процессов в нервных центрах возможно чередование сгибательных и разгибательных реакций конечностей, что необходимо для осуществления двигательного акта.
Конвергенция. Импульсы приходящие в ЦНС по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же промежуточным и эфекторным нейронам. Этот факт лег в основу принципа конвергенции, установленного Ч.Шеррингтоном. Конвергенция нервных импульсов объясняется тем, что на теле и дендритах каждого нейрона в ЦНС оканчиваются аксоны множества других нервных клеток. В спинном и продолговатом мозгу конвергенция имеет сравнительно ограниченный характер: на вставочных и моторных нейронах конвергируют афферентные импульсы, возникающие в различных участках рецептивного поля только одного и того же рефлекса. В отличие от этого в высших отделах ЦНС – в подкорковых ядрах и в коре больших полушарий – наблюдается конвергенция импульсов, исходящих из разных рецепторных зон. Поэтому один и тот же нейрон может возбуждаться импульсами, возникающими при раздражении и слуховых, и зрительных, и кожных рецепторов.
Принцип общего конечного пути. Этот принцип исходит из анатомического соотношения между афферентными и эфферентными нейронами. Количество чувствующих нейронов, приносящих возбуждение ЦНС, в 5 раз больше, чем двигательных. Соотношение между ними будет еще больше, если учесть, что вставочные нейроны являются воспринимающими нейронами в ЦНС. В связи с этим к одному мотонейрону приходит множество импульсов от различных рецепторов, но только некоторые из них приобретают рабочее значение. Таким образом, самые разнообразные стимулы могут быть причиной одной и той же рефлекторной реакции, т.е. происходит борьба за «общий конечный путь». Позднее было показано, что не количественное соотношение путей, а функциональные особенности нервных центров определяют, какой из множества нервных импульсов, сталкивающихся на пути к мотонейрону, окажется победителем и завладеет общим конечным путем. В ответ на множество различных раздражений всегда возникает биологически более значимая для организма реакция.
Принцип обратной связи. Воздействие работающего органа на состояние своего центра получило название обратной связи . Она обеспечивает длительное поддержание активности нервных центров, движение процессов возбуждения, торможения в ЦНС и зависит от постоянного притокавторичных афферентных импульсов. Импульсы, которые возникают в результате деятельности различных органов и тканей, называют, вторичными афферентными импульсами , а импульсы, идущие от рецепторов и вызывающие первичный рефлекторный акт, - первичными рефлекторными импульсами.
Вторичные афферентные импульсы возникают в мышцах, сухожилиях и суставах при осуществлении их деятельности. Они, постоянно поступая ото всех органов тела в ЦНС, способствуют ощущению положения нашего тела без зрительного контроля, обеспечивают поддержание нужного уровня функционирования нейронов в каждый данный момент.
Вторичная афферентная импульсация вносит постоянные поправки в осуществляющийся рефлекторный акт и обеспечивает наиболее тонкое приспособление организма к внешним воздействиям.
Афферентные импульсы, идущие от рабочих органов, способствуют созданиюаутогенного (собственного) торможения . Оно возникает в результате поступления в ЦНС афферентных импульсов от рецепторов – сухожильных рецепторов Гольджи. Эти рецепторы приходят в состояние возбуждения при растяжении или сокращении мышц. Возникший ТПСП уменьшает степень активности данного мотонейрона. Величина этих изменений может быть различной. Аутогенное торможение обеспечивает лучшее приспособление мышцы к осуществлению рефлекторного двигательного акта.
Факторы, обеспечивающие возможность координации:
1) Фактор структурно-функциональной связи – это наличие между отделами ЦНС, между ЦНС и различными органами функциональной связи, обеспечивающей преимущественное распространение возбуждения между ними. Прямая связь – управление другим центром или рабочим органом с помощью посылки к ни эфферентных импульсов, ПР: мозжечек посылает импульсы к ядрам ствола мозга. Обратная связь (обратная афферентация ) – управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них. Реципрокная связь – обеспечивает торможение центра – антагониста при возбуждении центра-агониста (мышцы сгибатели и разгибатели).
2) Фактор субординации – подчинение нижележащих отделов ЦНС вышележащим.
3) Фактор силы. Принцип общего конечного пути – в борьбе за общий конечный путь побеждает более сильное возбуждение (более важная команда в биологическом отношении), ПР: при слабом раздражении – рефлекс почесывания, при сильном – оборонительный рефлекс сгибание конечности, при одновременном раздражении возникает только оборонительный рефлекс).
4) Одностороннее проведение возбуждения в химических синапсах упорядочивает распространение возбуждения.
5) Феномен облегчения участвует при выработке навыков – возбуждение быстрее распространяется по проторенным путям, навыки становятся более координированными, ненужные движения постепенно устраняются.
6) Доминанта играет важную роль в процессах координации. Обеспечивает автоматизированное выполнение двигательных актов в процессе трудовой деятельности (доминанта двигательных центров).
Непрерывная смена процессов возбуждения и торможения в корковых клетках определяет цикличность работы отдельных органов и всего организма в целом. Этим объясняется иногда кажущаяся невероятная работоспособность некоторых выдающихся людей; недаром говорят, что гениальность на 90% заключается в высокой трудоспособности, которая во многом зависит от рациональной системы работы. Такую глубоко продуманную систему, как правило, создавали для себя все выдающиеся люди.
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов. Торможение в ЦНС – это активный процесс, внешне проявляющийся в подавлении, замедлении или ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью. Торможение нельзя свести ни к утомлению нервных центров, ни к их перевозбуждению, ни к катодической депрессии нервных клеток. Торможение в норме неразрывно связано с возбуждением, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному его распространению. При этом торможение часто ограничивает возбуждение и вместе с ним формирует мозаику активированных и заторможенных зон в центральных мозговых структурах.
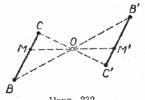
Явление торможения в нервной системе было открыто И.М.Сеченовым в 1862 году в виде эффекта центрального торможения. Суть его заключалась в том, что химическое раздражение кристалликом соли зрительных бугров (таламуса) лягушки вызывает торможение простых спинномозговых рефлекторных реакций.
Торможение – активный нервный процесс, приводящий к угнетению или предупреждению возбуждения. В зависимости от локализации тормозного процесса различают периферическое торможение, осуществляемое непосредственно в синапсах на мышечных и железистых элементах, и центральное, реализуемое в пределах центральной нервной системы.
Если торможение развивается как следствие возбуждения соответствующих дополнительных тормозящих или возбуждающих нейронов – это вторичное торможение.
Направленность эффекта (возбуждение или торможение) определяется изменением ионной проницаемости постсинаптической мембраны при взаимодействии медиаторов с рецепторами. Поэтому некоторые медиаторы способны опосредовать как возбуждение, так и Т.
У некоторых типов нейронов тормозные синапсы локализуются на телах и близких к ним участках Дендритов, что вследствие соседства с триггерной зоной генерации распространяющегося возбуждения обеспечивает высокую эффективность Т. Из этого правила есть исключения (например, тормозные синапсы звездчатых нейронов на клетках Пуркине мозжечка расположены на удалённых участках дендритов).
Функциональная значимость постсинаптического Т. разнообразна.
Афферентное (прямое) Т. служит для ослабления возбуждения функционально антагонистических элементов и тем самым способствует координированному, пространственно направленному протеканию возбуждения в цепях нейронов. В спинном мозге, в частности, такое Т. является основой так называемого реципрокного (взаимообратного) Т. мотонейронов, иннервирующих мышцы-антагонисты (см. Реципрокная иннервация). Возвратное (коллатеральное) Т., осуществляемое через систему возвратных коллатералей (ветвей) аксонов эфферентных нейронов и специализированных вставочных тормозных нейронов, стабилизирует собственный уровень возбуждения определённого структурно-функционального объединения (блока) нейронов и ограничивает распространение возбуждения на соседние популяции нейронов.
Менее изучено так называемое пресинаптическое Т., выражающееся в угнетении возбуждения в нервных терминалиях, то есть на входе постсинаптического клеточного элемента. Это Т. имеет необычайно большую длительность (сотни мсек ) и совпадает во времени с проявлением деполяризации приходящих афферентов. Предполагают, что на деполяризации основано пресинаптическое Т., а его морфологическим субстратом являются аксо-аксональные синапсы, происхождение пресинаптических элементов которых неизвестно. Имеются веские аргументы в пользу роли гамма-аминомасляной кислоты как медиатора пресинаптического Т., по крайней мере в нервно-мышечных соединениях ракообразных и в спинном мозге позвоночных. По-видимому, Сеченовское торможение у лягушки осуществляется по механизму преспнаптического Т.
Известно также пессимальное, или вторичное, Т., выражающееся в блокировании возбуждения вследствие его чрезмерности (см. Парабиоз). Этот феномен, описанный впервые Н. Е. Введенским (См. Введенский), трудно выявить при физиологических условиях эксперимента, но можно демонстрировать при аномальных (в частности, судорожных) состояниях.
Рис. Сеченовское торможение.
Торможение в ЦНС можно классифицировать по различным признакам:
по электрическому состоянию мембраны - деполяризационное и гиперполяризационное;
по отношению к синапсу - пресинаптическое и постсинаптическое;
по нейрональной организации – первичное и вторичное;
· по форме – латеральное (боковое), возвратное, реципрокное.


Рис. Проведение возбуждения от нейрона к нейрону. Локализация торможения – пресинаптическое и постсинаптическое. Пессимальное торможение
Первичное торможение обусловлено наличием специфических тормозных синапсов и развивается первично без предварительного возбуждения.
Вторичное торможение возникает без участия специализированных тормозных структур как следствие избыточной активации возбуждающих входов нейрона. Эту форму торможения определяют как торможение Введенского (пессимум), который открыл его в 1886 г. при исследовании нервно-мышечного препарата.
Торможение Введенского играет предохранительную роль и возникает при чрезмерной активации центральных нейронов в полисинаптических рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны, превышающей критический уровень и вызывающей инактивацию Na + –каналов, ответственных за генерацию потенциалов действия.
Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов импульсной активности нервных клеток.

Рис. Формы торможения в ЦНС.
В нервных центрах могут быть как различные виды торможения, так и разные их сочетания. В результате различают следующие формы торможения. Возвратное торможение заключается в том, что аксон тормозного нейрона, активированного возбуждающим нейроном, образует синапсы на той клетке, которая ее возбуждает, образуя своеобразную обратную связь тормозного типа. Возвратное торможение обнаружено в двигательных центрах передних рогов спинного мозга и служит, по-видимому, для дозирования возбуждения, приходящего к эфферентным нейронам.
В тех же двигательных центрах спинного мозга обнаружена еще одна форма торможения – реципрокное ((от лат. reciprocus - возвращающийся, обратный, взаимный) ) торможение, являющееся постсинаптическим. Такое торможение обеспечивает одновременное возбуждение нейронов мышц-сгибателей и торможение нейронов мышц-разгибателей на той же стороне тела, что необходимо для осуществления движения.

Рис. Эффект латерального торможения. Добавление тормозных вставочных нейронов сужает распределение частоты импульсации на высших уровнях.
Существует и еще одна форма торможения, получившая название латерального торможения, которое может быть как постсинаптическим, так и пресинаптическим. Возбуждающий нейрон через вставочные тормозные клетки вызывает торможение соседних нейронов, что приводит к формированию тормозной зоны, расположенной вокруг возбужденного нейрона. Латеральное торможение играет особо важную роль в сенсорных системах, при выделении наиболее важного признака стимула. Описанные формы торможения представлены на рисунке.
Торможение - особый нервный процесс, который обусловливается возбуждением и внешне проявляется угнетением другого возбуждения. Оно способно активно распространяться нервной клеткой и ее отростками. Основал учение о центральноv торможение И. М. Сеченов (1863), который заметил, что изгибающий рефлекс лягушки тормозится при химическом раздражении среднего мозга. Торможение играет важную роль в деятельности ЦНС, а именно: в координации рефлексов; в поведении человека и животных; в регуляции деятельности внутренних органов и систем; в осуществлении защитной функции нервных клеток.
Типы торможения в ЦНС
Центральное торможение распределяется по локализации на пре-и постсинаптическое;по характеру поляризации (зарядом мембраны) - на гипер-и деполяризации;
по строению тормозных нейронных цепей - на реципрокное, или соединенное, обратное и латеральное.
Пресинаптическое торможение , как свидетельствует название, локализуется в пресинаптических элементах и связано с угнетением проведения нервных импульсов в аксональных (пресинаптических) окончаниях. Гистологическим субстратом такого торможения является аксональные синапсы. К возбуждающему аксону подходит вставной тормозной аксон, который выделяет тормозной медиатор ГАМК . Этот медиатор действует на постсинаптическую мембрану, которая является мембраной возбуждающего аксона, и вызывает в ней деполяризацию. Возникшая деполяризация тормозит вход Са2 + из синаптической щели в заключение возбуждающего аксона и таким образом приводит к снижению выброса возбуждающего медиатора в синаптическую щель, торможение реакции. Пресинаптическое торможение достигает максимума через 15-20 мс и длится около 150 мс, то есть гораздо дольше, чем постсинаптическое торможение. Пресинаптическое торможение блокируется судорожными ядами - бикулином и пикротоксин, которые являются конкурентными антагонистами ГАМК .
Постсинаптическое торможение
(ГПСП) обусловлено выделением пресинаптическим окончанием аксона тормозного медиатора, который снижает или тормозит возбудимость мембран сомы и дендритов нервной клетки, с которой он контактирует. Оно связано с существованием тормозных нейронов, аксоны которых образуют на соме и дендритах клеток нервных окончаний, выделяя тормозные медиаторы - ГАМК и глицин . Под влиянием этих медиаторов возникает торможение возбуждающих нейронов. Примерами тормозных нейронов являются клетки Реншоу
в спинном мозге, нейроны грушевидные (клетки Пуркинье мозжечка), звездчатые клетки коры большого, мозга и др..
Исследованием П. Г. Костюка (1977) доказано, что постсинаптического торможения связано с первичной гиперполяризацией мембраны сомы нейрона, в основе которой лежит повышение проницаемости постсинаптической мембраны для К +. Вследствие гиперполяризации уровень мембранного потенциала удаляется от критического (порогового) уровня. То есть происходит его увеличение - гиперполяризация. Это приводит к торможению нейрона. Такой вид торможения называется гиперполяризационным.
Амплитуда и полярность ГПСП зависят от исходного уровня мембранного потенциала самого нейрона. Механизм этого явления связан с Сl + . С началом развития ТПСП Сl - входит в клетку. Когда в клетке становится его больше, чем снаружи, глицин конформирует мембрану и через открытые ее отверстия Сl + выходит из клетки. В ней уменьшается количество отрицательных зарядов, развивается деполяризация. Такой вид торможения называется деполяризационным.
Постсинаптическое торможение локальное.
Развивается оно градуально, способное к суммации, не оставляет после себя рефрактерности
. Является более оперативным, четко адресованным и универсальным тормозным механизмом. По своей сути это «центральное торможение», которое было описано в свое время Ch. S. Sherrington (1906).
В зависимости от структуры тормозного нейронного цепочки, различают следующие формы постсинаптического торможения: реципрокное, обратное и латеральное, которое является собственно разновидностью обратного.
Реципрокное (сочетанное) торможение характеризуется тем, что в том случае, когда при активизации афферентов возбуждаются, например, мотонейроны мышц-сгибателей, то одновременно (на этой стороне) тормозятся мотонейроны мышц-разгибателей, действующие на этот же сустав. Происходит это потому, что афференты от мышечных веретен образуют возбуждающие синапсы на мотонейронах мышц-агонистов, а через посредство вставного тормозного нейрона - тормозные синапсы на мотонейронах мышц-антагонистов. С физиологической точки зрения такое торможение очень выгодно, поскольку облегчает движение сустава «автоматически», без дополнительного произвольного или непроизвольного контроля.
Обратное торможение. В этом случае от аксонов мотонейрона отходит одна или несколько коллатералей, которые направляются в вставных тормозных нейронов, например, клеток Реншоу. В свою очередь, клетки Реншоу образуют тормозные синапсы на мотонейроны. В случае возбуждения мотонейрона активизируются и клетки Реншоу, вследствие чего происходит гиперполяризация мембраны мотонейрона и тормозится его деятельность. Чем больше возбуждается мотонейрон, тем больше ощутимые тормозные влияния через клетки Реншоу. Таким образом, обратное постсинаптическое торможение функционирует по принципу отрицательной обратной связи. Есть предположение, что этот вид торможения требуется для саморегуляции возбуждения нейронов, а также для предотвращения их перевозбуждению и судорожным реакциям.
Латеральное торможение. Тормозная цепь нейронов характеризуется тем, что вставные тормозные нейроны влияют не только на воспаленную клетку, но и на соседние нейроны, в которых возбуждение является слабым или вовсе отсутствует. Такое торможение называется латеральным, поскольку участок торможения, который образуется, содержится сбоку (латерально) от возбужденного нейрона. Оно играет особенно важную роль в сенсорных системах, создавая явление контраста.
Постсинаптическое торможения
преимущественно легко снимается при введении стрихнина, который конкурирует с тормозным медиатором (глицином) на постсинаптической мембране. Столбнячный токсин также подавляет постсинаптическое торможение, нарушая высвобождение медиатора из тормозных пресинаптических окончаний. Поэтому введение стрихнина или столбнячного токсина сопровождается судорогами, которые возникают вследствии резкого усиления процесса возбуждения в ЦНС, в частности, мотонейронов.
В связи с раскрытием ионных механизмов постсинаптического торможения появилась возможность и для объяснения механизма действия Вr. Натриq бромид в оптимальных дозах широко применяется в клинической практике как седативное (успокоительное) средство. Доказано, что такой эффект натрия бромида связан с усилением постсинаптического торможения в ЦНС. -
Роль различных видов центрального торможения
Главная роль центрального торможения заключается в том, чтобы во взаимодействии с центральным возбуждением обеспечивать возможность анализа и синтеза в ЦНС нервных сигналов, а следовательно, возможность согласования всех функций организма между собой и с окружающей средой. Эту роль центрального торможения называют координационной. Некоторые виды центрального торможения выполняют не только координационную, а и защитную (охранную) роль. Предполагают, что основная координационная роль пресинаптического торможения заключается в угнетении в ЦНС малосущественными афферентными сигналами. За счет прямого постсинаптического торможения согласуется деятельность антагонистических центров. Обратное торможение, ограничивая максимально возможную частоту разрядов мотонейронов спинного мозга, выполняет и координационную роль (согласовывает максимальную частоту разрядов мотонейронов со скоростью сокращения мышечных волокон, которые они иннервируют) и защитную (предотвращает возбуждению мотонейронов). У млекопитающих этот вид торможения распространен в основном в спинномозговых афферентных системах. В высших отделах мозга, а именно в корковом веществе большого мозга, доминирует постсинаптическое торможение.Какое функциональное значение пресинаптического торможения? За его счет осуществляется воздействие не только на собственный рефлекторный аппарат спинного мозга, но и на синаптические переключения ряда восходящих по головному мозгу трактов. Известно также о нисходящем пресинаптическом торможении первичных афферентных волокон группы Аа и кожных афферентов. В этом случае пресинаптическое торможение является, очевидно, первым «ярусом» активного ограничения информации, поступающей извне. В ЦНС, особенно в спинном мозге, пресинаптическое торможение часто выступает в роли своеобразной отрицательной обратной связи, которая ограничивает афферентную импульсацию при сильных (например, патологических) раздражениях и таким образом отчасти выполняет защитную функцию по отношению спинномозговых и выше расположенных центров.
Функциональные свойства синапсов не являются постоянными. В некоторых условиях эффективность их деятельности может расти или уменьшаться. Обычно при высоких частотах раздражения (несколько сот за 1 с) в течение нескольких секунд или даже минут облегчается синаптическая передача. Это явление получило название синаптической потенциации. Такая синаптическая потенциация может наблюдаться и по окончании тетанической стимуляции. Тогда она будет называться посттетанической потенциацией (ПТП). В основе ПТП (долговременного увеличения эффективности связи между нейронами), вполне вероятно, лежат изменения функциональных возможностей пресинаптического волокна, а именно его гиперполяризация. В свою очередь, это сопровождается повышением выхода медиатора в синаптическую щель и появлением увеличенного ВПСП в постсинаптической структуре. Есть данные и о структурных изменениях при ПТП (набухание и рост пресинаптических окончаний, сужение синаптической щели т.д.).
ПТП гораздо лучше выражена в высших отделах ЦНС (например, в гиппокампе, пирамидных нейронах коры большого мозга) по сравнению с спинномозговыми нейронами. Наряду с ПТП в синаптическом аппарате может возникать постактивационная депрессия, выражающаяся уменьшением амплитуды ВПСП. Эту депрессию многие исследователи связывают с ослаблением чувствительности к действию медиатора (десенсибилизации) постсинаптической мембраны или различным соотношением затрат и мобилизации медиатора.
С пластичностью синаптических процессов, в частности с ПТП, возможно, связаны формирования новых межнейронных связей в ЦНС и их закрепление, т.е. механизмы обучения и памяти. Вместе с тем следует признать, что пластические свойства центральных синапсов изучены пока недостаточно.
Координированная работа антагонистических нервных центров обеспечивается формированием реципрокных отношений между нервными центрами благодаря наличию специальных тормозных нейронов – клеток Реншоу.
Известно, что сгибание и разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Сигнал от афферентного звена через промежуточный нейрон вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель, а через клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель (и наоборот).
Латеральное торможение
При латеральном торможении возбуждение, передаваемое через коллатерали аксона возбужденной нервной клетки, активирует вставочные тормозные нейроны, которые тормозят активность соседних нейронов, в которых возбуждение отсутствует или является более слабым.
В результате в этих соседних клетках развивается очень глубокое торможение. Образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону.
Латеральное торможение по нейронному механизму действия может иметь форму как постсинаптического, так и пресинаптического торможения. Играет важную роль при выделении признака в сенсорных системах, коре больших полушарий.
Значение торможения
Координация рефлекторных актов . Направляет возбуждение к определенным нервным центрам или по определенному пути, выключая те нейроны и пути, деятельность которых в данный момент является несущественной. Результатом такой координации является определенная приспособительная реакция.
Ограничение иррадиации .
Охранительное. Предохраняет нервные клетки от перевозбуждения и истощения. Особенно при действии сверхсильных и длительно действующих раздражителей.
В реализации информационно-управляющей функции ЦНС значительная роль принадлежит процессам координации деятельности отдельных нервных клеток и нервных центров.
Координация – морфофункциональное взаимодействие нервных центров, направленное на осуществление определенного рефлекса или регуляции функции.
Морфологическая основа координации: связь между нервными центрами (конвергенция, дивергенция, циркуляция).
Функциональная основа: возбуждение и торможение.
Основные принципы координационного взаимодействия
Сопряженное (реципрокное) торможение .
Обратная связь .Положительная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в том же направлении, что и основные сигналы, что ведет к усилению рассогласования в системе.Отрицательная – сигналы, поступающие на вход системы по цепи обратной связи, действуют в противоположном направлении и направлены на ликвидацию рассогласования, т.е. отклонений параметров от заданной программы (П.К. Анохин).
Общий конечный путь (принцип «воронки»Шеррингтона ). Конвергенция нервных сигналов на уровне эфферентного звена рефлекторной дуги определяет физиологический механизм принципа «общего конечного пути».
Облегчение .Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов выше суммы реакций при изолированном раздражении этих рецептивных полей.
Окклюзия . Это интегративное взаимодействие нервных центров, при котором суммарная реакция при одновременном раздражении рецептивных полей двух рефлексов меньше, чем сумма реакций при изолированном раздражении каждого из рецептивных полей.
Доминанта .Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в ЦНС. ПоА.А. Ухтомскому , доминантный очаг характеризуется:
Повышенной возбудимостью,
Стойкостью и инертностью возбуждения,
Повышенной суммацией возбуждения.
Доминирующее значение такого очага определяет его угнетающее влияние на другие соседние очаги возбуждения. Принцип доминанты определяет формирование главенствующего возбужденного нервного центра в тесном соответствии с ведущими мотивами, потребностями организма в конкретный момент времени.
7. Субординация. Восходящие влияния преимущественно носят возбуждающий стимулирующий характер, нисходящие носят угнетающий тормозной характер. Эта схема согласуется с представлениями о росте в процессе эволюции роли и значении тормозных процессов в осуществлении сложных интегративных рефлекторных реакций. Имеет регулирующий характер.